こんにちは!
それでは今回も化学のお話やっていきます。
テーマはこちら!
動画はこちら↓

動画で使ったシートはこちら(polymer distribution)
合成高分子と生体高分子
まずは、化学的な合成によってできる高分子と生体由来の高分子の違いについて、見ていきましょう。

例
合成高分子の例は、ポリエチレンやポリスチレンなどがあります。
ポリエチレンは買い物袋やお弁当のフタ、ポリスチレンは言うまでもなく発泡スチロールの素材です。
あとは、CDやDVDのケースもポリスチレンです。
高分子の名前は、アルファベットで略して表記することが多く、ポリエチレンならPE、ポリスチレンならPSと表しますので、ぜひ知っておいてください。
一方で、DNAやRNAがある核酸とコラーゲンやアミラーゼなどのタンパク質、デンプンやセルロースなどの多糖は、生体高分子に分類されます。
それぞれの構成要素はヌクレオチド、アミノ酸、単糖になります。
分子量
合成高分子と生体高分子の違いとして、まず分子量の分布が挙げられます。
簡単なのは生体高分子で、基本的に単分散、すなわちある特定の分子量をもつものしかありません。
それに対して、合成高分子は分子量に分布があり、完全な単分散の試料を得ることはできません。
つまり、分子量10万の高分子を作ろうとしても、10万5000や9万5000など近い大きさのものも混ざったものしか得ることができないということです。
その理由は、分子量と物理的性質の関係にあります。
例えば、メタンとエタンのように低分子の場合は、沸点や溶解度にある程度の違いがあります。
しかし、大きな分子になってくると、それが1000量体だろうと1001量体だろうと物性はほとんど変わらなくなっていきます。
その結果、特定の大きさの分子だけを単離することができず、混合物になります。
合成によって純粋な物質が得られないというのが化学(chemistry)と高分子科学(polymer science)で大きく違うことで、当初はそこが化学者の反感を買いました。
高分子の分子量を議論する場合には、平均の分子量と分布の広がりを考えます。
ただし、今日では分子量分布の狭い高分子を作るリビング重合という合成法も確立されてきており、物性の研究もしやすくなっています。
配列制御
続いての違いは、モノマー配列です。
具体的には、2種類以上のモノマーがあるときに見られる並び方の法則に違いがあります。
生体高分子の場合、デンプンやセルロースなどの単純な多糖を除いて、核酸やタンパク質ではその配列がほぼ完全に制御されています。
核酸の鋳型がすでに存在していて、その情報から分子が作られます。
その過程でたまにミスも起こりますが、触媒となるタンパク質、すなわち酵素には修正する機能をもつものがあるため、最終的にミスが残ることはほとんどありません。
残念ながら、ミスが取り除けなかった場合は、それががんなどの遺伝子の異状や突然変異につながります。
対して、合成高分子ではここまでの配列制御はできておらず、ざっくりとしか制御することはできません。
分子の形状
次の違いは、分子の形状です。
どちらでも線状、すなわち分岐がない一本の高分子が多く見られますが、もちろんそれだけではなく、生体高分子では日本のお米に多く含まれるアミロペクチンのような枝分かれ構造や原核生物の環状DNAなども見られます。
特に、線状のタンパク質の一部は、分子内・分子間相互作用により、特異的な立体構造を有することで、触媒として作用するようになります。
合成高分子においては、そのほとんどが線状か、ゴムに代表される高分子網目構造です。
一部例外として、レジ袋に使われる低密度ポリエチレンは枝分かれ構造をもっているほか、環状高分子の合成例が少しあります。
高分子が担う役割
最後、高分子が担う役割に違いがあります。
ここに挙げたものがすべてではないですが、まず合成高分子の役割は衝撃吸収、軽量化、接着、絶縁、コーティングなどがあります。
高分子はエネルギーを熱として分散させることができ、その軽い質量の割に耐衝撃性や引張強度が大きいという特徴があります。
これにより、自動車や飛行機の部品が金属から高分子に変わってきています。
そして、生体高分子には、合成高分子にはないユニークなものがたくさんあります。
触媒、情報の保存と伝達、細胞や組織の骨格維持、細胞認識に用いるための目印、そしてオートファジーというのが有名ですが、いざというときには分解することでエネルギーに変えることができます。
特に特徴的なのは、やはり分子に情報を書き込むことができる点で、高い精度での配列制御によってそれが可能になっています。
単独重合体と共重合体
続いて、ほかの高分子の分類法について見ていきます。
紹介するのは、単独重合体と共重合体です。
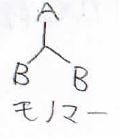
高分子を構成する最小単位が1種類しかないとき、それを単独重合体と呼び、2種類以上のモノマーからできるものを共重合体と呼びます。
先述のとおり、合成高分子の配列制御は生体高分子に比べてかなり大雑把であり、その並び方の特徴から、さらに下記のように分類されます。

ここでは2種類のモノマーを\(\displaystyle \rm{A}\)、\(\displaystyle \rm{B}\)と表すことにしています。
\(\displaystyle \rm{ABAB}\)というように、交互に違うモノマーが連なるものは交互共重合体と呼びます。
そして\(\displaystyle \rm{AAAABBBB}\)というように、Aが連続した部分とBが連続した部分があるものはブロック共重合体と言います。
\(\displaystyle \rm{A}\)が3個つながったら\(\displaystyle \rm{B}\)が1個、そしてまた\(\displaystyle \rm{A}\)が3個というように、周期的な配列をとる場合は周期共重合体と呼びます。
法則がなく、ばらばらにつながっているものはランダム共重合体と言います。
生体高分子である核酸やタンパク質も共重合体の一種であり、それぞれ4種類のヌクレオチドと20種類のアミノ酸の共重合体になります。
結合様式による分類
最後に、結合様式から見た分類法を見ていきます。
ここでは、モノマーの化学的な構造には注目せずに分子の形にのみ注目します。
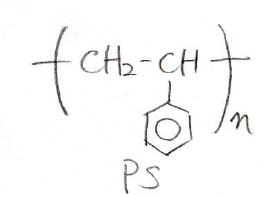
まず1つ目が最もシンプルな線状高分子です。

身の回りで言うと、PETボトルでおなじみのポリエチレンテレフタレートや発泡スチロールのポリスチレンが該当します。
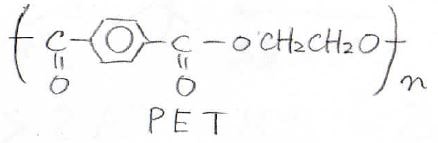
そして、別の形状として環状高分子があります。

先述のとおり、合成で作るのは難しいとされています。
これは、環を作るためには一度長い分子を作った後に両端を反応させる必要がありますが、長くなるほど両端が出会う確率が下がるためです。

生体高分子としては、大腸菌などの原核生物やミトコンドリアなどのDNAが環状になっていますが、真核生物のDNAがなぜ線状分子になったのかというのは遺伝学でいろいろな説が提唱されています。
より高度な生物となるためには大きなDNAが必要になるので、それを細胞核に収納するためにはタンパク質に巻き付けやすい線状がよかった説や、ミトコンドリアやウイルスのDNAを自らに取り込んで進化するために線状のほうが都合がよかった説などがあります。
話を戻して、次に紹介する形状は、分岐高分子です。
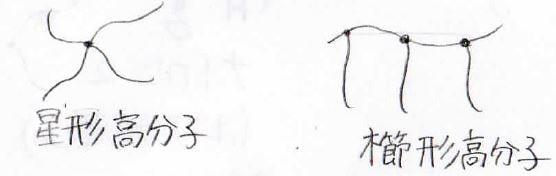
分岐高分子はさらに分岐の仕方によって分類があって、まず分子内で1つの分岐点しかもたないものは星形高分子と呼びます。
そして、一本の鎖だけにいくつもの分岐点をもつものは櫛形高分子と言います。
下図のように、主鎖には\(\displaystyle \rm{A}\)が連続していて、そこに\(\displaystyle \rm{B}\)が連続した鎖が何本も付いた櫛形の共重合体は、グラフト共重合体と呼びます。
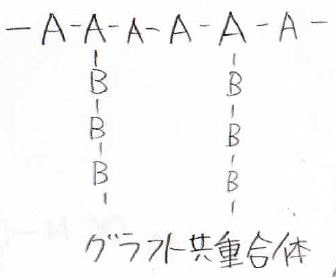
また、別の分岐高分子として、ランダム分岐高分子のほかに、高度に架橋点が密集したハイパーブランチポリマーというものがあります。



それと似て非なるものとして、中心から規則正しく高度な分岐構造をもつ高分子のことは、デンドリマーと言います。

上図は、モノマーが3官能基型で、\(\displaystyle \rm{AB}\)間で結合が形成されるとしてできるデンドリマーです。
モノマーの中に分岐点があるので、分岐点間の長さはすべて同じになります。
そして、これが形状での分類の最後です。
共有結合によって網目を形成した網目状高分子です。

最も一般的なのは、天然ゴムに硫黄を反応させて架橋させたゴムです。
この網目は全体でつながっているので、とても巨大な1つの分子だということになります。
輪ゴムも1個で1つの分子です。
ここまで大きくなると、もはや分子量による物理的な性質の違いは生まれず、架橋点を結んだその間の長さの平均やその分布、鎖の剛直性や可塑剤として混ぜられる低分子の存在によってその力学特性が変わることになります。
そして、網目状高分子の少し変わったパターンとして、2種類以上の異なる網目状高分子が互いに組み込まれて外れなくなった、相互侵入高分子網目、通称IPNと呼ばれる系があります。

一方だけをある反応で分解したり、会合系ネットワークにすることで材料の力学特性を複雑にコントロールすることが可能になります。
接着剤や増粘剤などで研究が盛んにされている分野です。
まとめ
今回の内容は以上です。
間違いの指摘、リクエスト、質問等あれば、Twitter(https://twitter.com/bakeneko_chem)かお問い合わせフォームよりコメントしてくださると、助かります。
それではどうもありがとうございました!